
Содержание:
- 1. Свойства генов
- 2. Взаимодействие аллельных генов
- 3. Взаимодействие неаллельных генов
- 4. Сцепление генов. Опыты Моргана
- 5. Кроссинговер
- 6. Хромосомная теория наследственности
- 7. Генетические карты
- 8. Цитоплазматическая наследственность
Свойства генов
♦ Свойства генов и особенности их проявления в признаках:
■ ген дискретен в своем действии, т.е. обособлен в своей активности от других генов;
■ один ген отвечает за проявление одного строго определенного признака или нескольких признаков (плейотропия);
■ один признак может быть результатом действия нескольких генов (аллельных или неаллельных);
■ ген может усиливать степень проявления признака при увеличении числа его доминантных аллелей;
■ ген может взаимодействовать с другими генами; это приводит к появлению новых признаков;
■ изменение положения гена в хромосоме или влияние факторов внешней среды могут модифицировать его проявление в признаках;
■ ген обладает способностью к мутациям.
Взаимодействие аллельных генов
Взаимодействие генов — явление, когда за один признак отвечает несколько генов (или аллелей).
Аллельное взаимодействие — это взаимодействие аллелей одного и того же гена (за признак отвечает несколько аллелей одного и того же гена).
❖ Типы аллельных взаимодействий:
■ доминирование,
■ неполное доминирование,
■ сверхдоминирование,
■ кодоминирование.
Доминирование — тип взаимодействия двух аллелей одного гена, когда один (доминантный) из них полностью исключает действие другого (рецессивного). Примеры: доминирование у человека темных волос над светлыми, карих глаз над голубыми.
Неполное доминирование — степень активности доминантного аллеля недостаточна для того, чтобы полностью подавить действие рецессивного аллеля и обеспечить полное проявление доминантного признака.
■ В этом случае у гетерозигот формируется промежуточный (по отношению к родительским признакам) признак — имеет место промежуточный характер наследования. Этот признак будет наблюдаться у гибридов первого поколения и гетерозигот второго поколения. Во втором поколении расщепление по фенотипу и генотипу оказывается одинаковым 1:2:1 (одну часть составляет доминантная гомозигота АА с выраженным доминантным признаком, две части составляет гетерозигота Аа с промежуточным признаком и одну часть составляет гомозигота аа с рецессивным признаком).
■ Примеры неполного доминирования: наследование формы (курчавости) волос у человека, масти крупного рогатого скота, окраски цветков у растения ночная красавица (см. таблицу).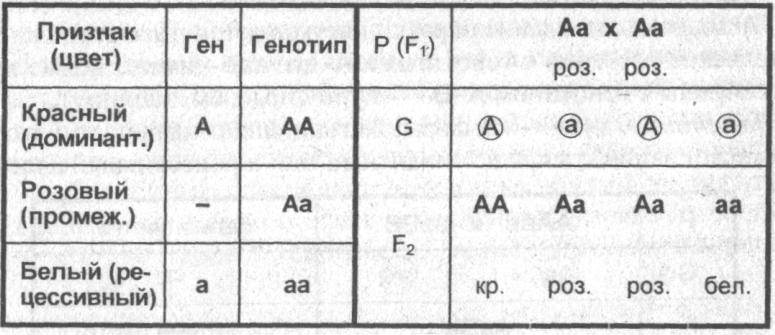
— Регулярная проверка качества ссылок по более чем 100 показателям и ежедневный пересчет показателей качества проекта.
— Все известные форматы ссылок: арендные ссылки, вечные ссылки, публикации (упоминания, мнения, отзывы, статьи, пресс-релизы).
— SeoHammer покажет, где рост или падение, а также запросы, на которые нужно обратить внимание.
SeoHammer еще предоставляет технологию Буст, она ускоряет продвижение в десятки раз, а первые результаты появляются уже в течение первых 7 дней. Зарегистрироваться и Начать продвижение
Сверхдоминирование — более сильное проявление признака у гетерозиготной особи (Аа), чем у любой из гомозигот (АА и аа).
Кодоминирование — оба аллеля равноценны, не подавляют друг друга и участвуют в определении признака у гетерозиготной особи. Пример: наследование IV группы крови у человека, которая детерминируется одновременным присутствием в генотипе двух кодоминантных генов IА и Iв. Первый из этих генов детерминирует синтез в эритроцитах белка-антигена А, второй — синтез белка-антигена В; наличие обоих этих генов в генотипе приводит к тому, что у людей с IV группой крови эритроциты содержат как белок-антиген А, так и белок-антиген В.
Взаимодействие неаллельных генов
Неаллельное (или межаллельное) взаимодействие — это взаимодействие аллелей разных генов, т.е. генов, располагающихся в негомологичных хромосомах или разных локусах гомологичных хромосом.
■ Неаллельное взаимодействие генов приводит к модификации менделевского расщепления по фенотипу 9 : 3 : 1, т.е. к появлению в потомстве гетерозиготы иных расщеплений, например 9 : 3 : 4; 9 : 6 : 1; 12 : 3 : 1 и др.
❖ Основные типы межаллельных взаимодействий:
■ комплементарность;
■ эпистаз;
■ полимерия.
Замечание: комплементарное и эпистатическое взаимодействия возникают в тех случаях, когда признак контролируется одной парой неаллельных генов.
Комплементарное, или дополнительное, взаимодействие — такой тип межаллельного взаимодействия генов, при котором одновременное присутствие в генотипе гибрида доминантных генов разных аллельных пар приводит к появлению нового признака, отсутствующего у обоих родителей.
Пример: наследование окраски цветков душистого горошка (родительские растения с генотипами А-вв, ааВ- имеют белые цветки, гибриды с генотипом А-В- пурпурные; см. таблицу).
Замечание: знак «-» в формуле генотипа означает, что это место может занимать как доминантный, так и рецессивный аллель.
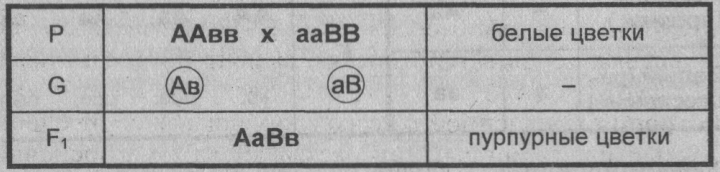
Объяснение: пурпурный пигмент образуется с помощью специального фермента, который синтезируется только при наличии обоих доминантных генов: как А, так и В. Цветки родительских особей имеют белый цвет, так как в генотипе каждого из них присутствует только один из этих генов.
Во втором поколении при самоопылении, обеспечивающем равновероятное (случайное) образование гамет и зигот разного типа, наблюдается расщепление по фенотипу в соотношении пурпурной и белой окраски цветков как 9 : 7 (9 пурпурных: A-В- и 7 белых: ЗА-Bb, ЗааВ-. 1aabb).
— Разгрузит мастера, специалиста или компанию;
— Позволит гибко управлять расписанием и загрузкой;
— Разошлет оповещения о новых услугах или акциях;
— Позволит принять оплату на карту/кошелек/счет;
— Позволит записываться на групповые и персональные посещения;
— Поможет получить от клиента отзывы о визите к вам;
— Включает в себя сервис чаевых.
Для новых пользователей первый месяц бесплатно. Зарегистрироваться в сервисе
Эпистаз — такой тип межаллельного взаимодействия генов, при котором аллели одного гена подавляют проявление аллельной пары другого гена, и подавляемый признак не проявляется.
Супрессор (или ген-ингибитор) — ген, подавляющий действие других неаллельных генов. Супрессором может быть как доминантный, так и рецессивный ген.
Доминантный эпистаз — эпистаз, в котором супрессором является доминантный ген. При доминантном эпистазе во втором поколении наблюдается расщепление фенотипа 12:3:1 или 13:3.
Рецессивный эпистаз (криптомерия) — эпистаз, в котором супрессором является рецессивный ген. При рецессивном эпистазе во втором поколении наблюдается расщепление по фенотипу 9:3:4.
Пример эпистаза: наследование окраски шерсти у домашних кроликов. Синтез черного пигмента детерминирует рецессивный ген с, доминантная аллель I другого гена является супрессором, подавляя действие гена с. Тогда кролики с генотипами C-I-, ccl-будут белыми, кролики с генотипами C-ii — серыми, а с генотипом ссii — черными.
Многие признаки контролируются двумя и более парами неаллельных генов (называемых в этом случае полимерными).
Полимерия — взаимодействие нескольких неаллельных полимерных генов. При полимерии степень выраженности фенотипического признака часто зависит от числа полимерных генов, ответственных за его проявление. При кумулятивной полимерии действие генов суммируется; примеры: масса тела, молочность крупного рогатого скота, яйценоскость кур, некоторые параметры умственных способностей человека и др. При некумулятивной полимерии степень проявления признака от числа доминантных генов в генотипе не зависит (пример: оперенность ног у кур).
Плейотропия — зависимость нескольких признаков от одного гена. Каждый плейотропный ген оказывает какое-то основное действие, но модифицирует проявление других генов.
Сцепление генов. Опыты Моргана
Сцепленные гены — любые гены, расположенные в одной хромосоме.
Группа сцепления — все гены, расположенные в одной хромосоме.
■ Количество групп сцепления равно числу пар хромосом (т.е. гаплоидному числу хромосом). У человека 46 хромосом, т.е. 23 группы сцепления.
■ Наследование признаков, за которые отвечают гены из одной группы сцепления, не подчиняется законам Менделя.
Опыты Т. Моргана (1911-1912 гг.): анализ наследования двух пар альтернативных признаков у мух-дрозофил — серого (В) и черного (b) цвета тела и нормальной (V) или укороченной (v) длины крыльев.
Первая серия опытов: скрещивание гомозиготной доминантной (BBVV) особи (с серым цветом тела и нормальной длиной крыльев) с гомозиготной рецессивной (bbvv) особью черного цвета с короткими крыльями. Все потомки F1, в соответствии с первым законом Менделя, являются доминантными гетерозиготными (BbVv) особями серого цвета с нормальными крыльями.
Вторая серия опытов: анализирующее скрещивание гибридов первого поколения — гомозиготной рецессивной (черной короткокрылой) самки (bbvv) с дигетерозиготным (серым с нормальными крыльями) самцом (BbVv). Если предположить, что два гена, относящиеся к разным аллельным парам, локализованы в разных хромосомах, то у дигетерозиготы следует ожидать образования (в равных количествах) четырех типов гамет: BV, bV, Bv и bv. Тогда, согласно третьему закону Менделя, в потомстве должны присутствовать четыре разных фенотипа в равном количестве (по 25%). В действительности присутствовали только два фенотипа (в соотношении 1 : 1).
■ Это означает, что доминантные гены В и V, относящиеся к разным аллельным парам, локализованы в одной хромосоме (из пары гомологичных хромосом) и попадают в одну гамету, а оба рецессивных гена в и v локализованы в другой хромосоме и вместе попадают в другую гамету. Поэтому у дигетерозиготного самца мухи-дрозофилы образуется не четыре типа гамет (когда гены расположены в разных хромосомах), а только два: BV (50%) и bv (50%), и, следовательно, потомки F2 будут иметь два сочетания признаков.
Третья серия опытов: проверка предположения о полном сцеплении генов путем анализирующего скрещивания дигетерозиготной (серой с нормальными крыльями) самки (BbVv) из поколения F1 с гомозиготным рецессивным (черным короткокрылым) самцом (bbvv) из родительского поколения. В результате были получены потомки четырех фенотипов в следующем соотношении: по 41,5% особей с серым телом и нормальными крыльями (генотип BbVv) и особей с черным телом и короткими крыльями (генотип bbvv), и по 8,5% серых короткокрылых особей (генотип Bbvv) и черных особей с нормальными крыльями (генотип bbVv).
Отсюда следует, что сцепленные гены, т.е. гены, локализованные в одной хромосоме, не всегда передаются вместе, т.е. сцепление может быть неполным. Это связано с явлением кроссинговера. вероятность которого в данном случае составляет 17%.
Кроссинговер
Кроссинговер — явление обмена участками гомологичных хрома-тид в процессе их конъюгации в профазе мейоза I.
■ У гетерозиготных организмов кроссинговер приводит к пере-комбинации генетического материала.
■ Кроссинговер происходит не всегда; его частота зависит от расстояния между генами (подробнее о расстоянии между генами см. ниже).
■ Кроссинговер имеет место у всех растений и животных, за исключением самца мухи-дрозофилы и самки тутового шелкопряда.
■ Значение кроссинговера: он позволяет создавать новые комбинации генов и тем самым повысить наследственную изменчивость, необходимую для расширения возможностей адаптации организмов к изменившимся условиям среды.
Рекомбинация — появление новых сочетаний генов в результате кроссинговера, свободного комбинирования хромосом при образовании гамет или при их слиянии в ходе оплодотворения.
Кроссоверные (или рекомбинантные) особи — особи, образующиеся из гамет с новым, полученным путем кроссинговера, сочетанием аллелей.
Закон Моргана (закон сцепленного наследования): гены, расположенные в одной хромосоме (т.е. входящие в одну группу сцепления), наследуются преимущественно, т.е. с наибольшей вероятностью, вместе (сцепленно).
Хромосомная теория наследственности
Хромосомная теории наследственности экспериментально обоснована Т. Морганом с сотрудниками в 1911-1926 гг.
♦ Основные положения хромосомной теории наследственности:
■ гены, ответственные за хранение и передачу наследственной информации, локализованы в определенных участках (локусах) хромосом; различные хромосомы имеют разное число генов;
■ аллельные гены занимают одинаковые локусы в гомологичных хромосомах;
■ в хромосомах гены располагаются в линейной последовательности (друг за другом) и не перекрываются;
■ гены гомологичных хромосом образуют группу сцепления и наследуются преимущественно вместе; число групп сцепления равно гаплоидному набору хромосом (т.е. числу пар гомологичных хромосом);
■ между гомологичными хромосомами возможен обмен аллельными генами (кроссинговер);
■ вероятность кроссинговера пропорциональна расстоянию между генами в хромосомах.
Генетические карты
Генетическая карта хромосом — схема, на которой отображается взаимное расположение генов, находящихся в одной группе сцепления, с учетом их порядка следования и относительных расстояний между ними.
■ Возможность картирования хромосом основана на постоянстве процента кроссинговера между определенными генами.
Расстояние между генами выражается в морганидах.
■ Одна морганида — такое расстояние между генами, при котором вероятность кроссинговера равна 1 %.
♦ Значение генетических карт:
■ в селекции они позволяют прогнозировать возможность получения организмов с определенными сочетаниями признаков;
■ являются основой для генной инженерии;
■ в медицине используются для диагностики ряда тяжелых наследственных заболеваний человека;
■ сравнение генетических карт разных видов живых организмов помогает установить особенности эволюционного процесса.
Цитоплазматическая наследственность
Цитоплазматическая наследственность — наследственность, связанная с действием генов, находящихся в органоидах цитоплазмы, содержащих ДНК (митохондриях и пластидах).
■ Такие гены способны к автономной репликации и равномерному распределению между дочерними клетками.
■ В передаче признаков цитоплазматическая наследственность имеет второстепенное значение.
■ Цитоплазматическое наследование осуществляется только через материнский организм (в мужских половых клетках цитоплазмы мало и в ней митохондрии и пластиды отсутствуют). Примеры: с мутациями генов, локализованных в митохондриях, связано наследование нарушений в действии дыхательных ферментов у дрожжей; с мутациями генов, локализованных в пластидах, связано, в частности, наследование пестролистности у ряда растений (ночной красавицы, львиного зева и др.).

Написать ответ